O slepičích zubech a ještěřím peří: embrya, geny a evoluce
Pokud by evoluční změny skutečně vyplynuly z disociace indukčních událostí, mohli bychom očekávat, že zbytky starých vztahů přetrvávají, že regulační geny, které nebyly exprimovány stejným způsobem jako dříve, by stále byly přítomny. Toto očekávání je opodstatněné, jak ukazuje obr. 5-6, na kterém O. Mangold schematicky znázornil některé z hlavních indukčních událostí v hlavě a trupu embryí obojživelníků. Tento diagram zdůrazňuje obecný závěr, který již byl vyvozen z Jacobsonovy práce, o složitosti indukčních interakcí, které jsou obvykle propleteného nebo kaskádového typu. Aplikováno na evoluci to znamená, že indukční interakce jsou odolné vůči velkým modifikacím a že když dojde k dostatečně velkým změnám, obvykle se nacházejí v mezích charakteristických pro danou skupinu. Radikální změny, které jsou spojeny se vznikem některých nových skupin, jako jsou členovci (viz kapitola 4), se někdy vyskytují, ale je to vzácné. Dalším často zmiňovaným důsledkem integrované povahy morfogenetických drah je, že evoluční změny v pozdních stádiích vývoje jsou snadněji vnímatelné než změny rané, jednoduše proto, že pozdní změny vyžadují méně odpovídajících změn v kaskádových procesech, které ovlivňují. Je však stěží možné předpokládat, že změny v ontogenezi vznikají výhradně tímto způsobem; pokud by to byla jediná cesta, něco velmi blízkého Haeckelovské rekapitulaci by skutečně hrálo univerzální roli. Změny ve skutečnosti vznikají ve všech fázích vývoje, takže ačkoli vzorce interakcí pozorované u obojživelníků lze považovat za jakýsi archetyp pro vývoj obratlovců, žádné dva druhy obratlovců se nevyvíjejí úplně stejně, a to ani v jejich nejranějších fázích. Raná podobnost a následné rostoucí oddělování embryí příbuzných organismů, o nichž se hovoří v Baerových zákonech, by neměly být brány příliš doslova. Vyvíjející se savci nejsou v žádné fázi vývoje identické s embryi ryb nebo ještěrek: ve svých genomech a vývojových vzorcích se od svých předků velmi vzdálili. Přesto se mezi nimi zachovaly některé prvotní podobnosti. Stalo se to, že mnohočetné indukční interakce, popsané v příkladech indukce čočky, nosu a ucha, vytvořily dostatečně vysokou úroveň morfogenetické homeostázy, takže změny lze vnímat i v raných fázích celkového vývojového procesu, aniž by byla narušena jeho integrace.
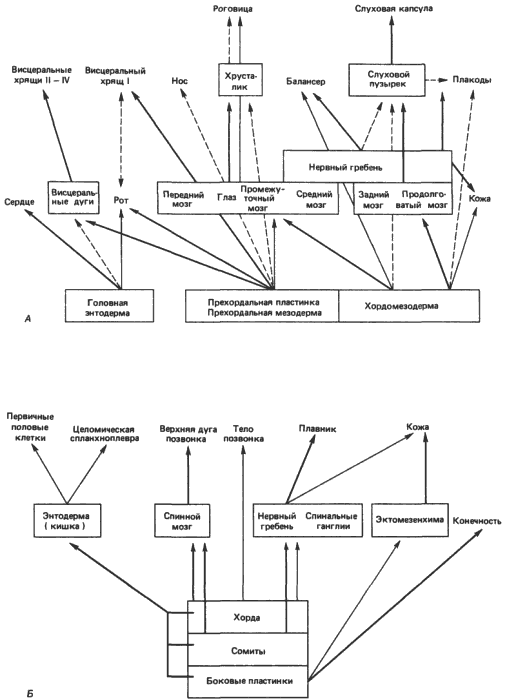
Rýže. 5-6. Indukční interakce a kaskádové procesy ve vývoji obojživelníků. A. Indukční procesy ve vývoji hlavy. B. Indukční procesy ve vývoji kmene. Tloušťka šipek odráží relativní intenzitu indukce (Mangold, 1961).
Existence obecného konzervativního vývojového plánu naznačuje, že embryonální struktury mohou být zachovány i poté, co ztratily svou předchozí základní funkci, protože stále slouží jako spojovací články v kaskádě morfogenetických interakcí, které jsou s předchozí funkcí inherentní. Regulační systémy mohou být zachovány stejným způsobem. Důkazem zachování předchozích regulačních genů by mohlo být vzkříšení dříve odmítnuté vývojové dráhy, když je stávající typ vývoje narušen mutací nebo experimentálním vlivem. Příklady obou jsou známé. Obnova starých typů vývoje v důsledku mutací je podrobně diskutována v kapitolách 8 a 9. Experimentální analýza tří příkladů evolučních změn odhalila modifikace v systémech interakce mezi tkáněmi, ve kterých byly nové regulační mechanismy superponovány na předchozí, ale bez ztráty genetické informace přítomné u předků. Dva z těchto příkladů se vztahují k modifikacím, ke kterým došlo v evoluci ptáků; třetí se týká změn v kožních tkáních plazů, ptáků a savců.
Archaeopteryx je již dlouho známý jako „chybějící článek“, protože měl ptačí rysy, jako je peří, nepatřičně kombinované s plazí, nebo přesněji dinosauří kostrou, a to až po dlouhý ocas a zuby. Mnoho změn, které se u ptáků vyvinuly, vedlo ke zvýšení efektivity letu díky úbytku hmotnosti, což bylo doprovázeno ztrátou některých charakteristických rysů Archaeopteryxe, včetně zubů, drápů na předních končetinách a dlouhého plazího ocasu, stejně jako redukcí nebo srůstem některých kostí končetin. Poslední ozubení ptáci pocházejí z pozdní křídy, tedy dostatečně dlouho na to, aby ptačí genomy byly zbaveny genetické informace potřebné pro morfogenezi zubů. Ve skutečnosti se tak nestalo a je skutečně možné vyvolat zuby u kuřat.
Pro normální proces vývoje zubu, jak jej shrnuli Thesleff a Deuchar, jsou nezbytné reciproční indukční interakce mezi ústním epitelem a podkladovým ústním mezenchymem. Sled událostí, které k tomu dochází, je schematicky znázorněn na obr. 5-7. Ústní mezenchym indukuje z nadložního epitelu vývoj sklovinného orgánu; tento orgán následně invaginuje do mezenchymu a indukuje jeho diferenciaci na odontoblasty, které vylučují dentin. Při absenci sklovinného orgánu dává ústní mezenchym vzniknout pouze houbovité kosti. Do vývoje zubu se zapojuje i další indukční proces: epitelové buňky vnitřní části sklovinného orgánu reagují na přítomnost mezenchymálních odontoblastů diferenciací na ameloblasty, které vylučují zubní sklovinu.
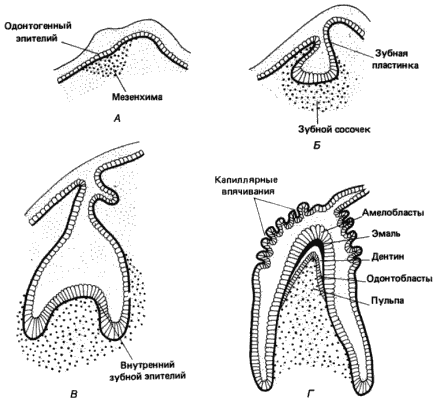
Rýže. 5-7. Morfogeneze zubů u savců. A. Vrstva epiteliálních buněk zubní laminy, která leží nad ústním mezenchymem. B. Epitelové buňky pod vlivem indukčního signálu z mezenchymu zaplavují tento tkáň. V. Zubní ploténka, indukovaná k vytvoření sklovinového orgánu, následně indukuje diferenciaci mezenchymu do odontoblastů. G. Vznik zubu v důsledku ukládání dentinu mezenchymálními odontoblasty a skloviny epiteliálními odontoblasty.
U ptáků vzniká v důsledku normální interakce mezi ústním mezenchymem a epidermis zobák, nikoli zuby. Hayashi s využitím heterospecifických kombinací ústních tkání kuřat a kachen ukázal, že charakter zobáku je určen druhem mezenchymu. Když byl tedy embryonální epitel kuřat zkombinován s embryonálním mezenchymem kachny, vyvinul se zobák s charakteristickými dentiklemi kachny. Podobně Kollar a Baird zjistili, že myší ústní epitel je ovlivněn mezenchymem, se kterým je vyrostl.
Nejodhalujícím „evolučním“ experimentem je kombinace ptačího ústního epitelu s mezenchymem savčí ústní dutiny. Collard a Fischer provedli tento experiment pěstováním kousků kuřecího faryngeálního epitelu s myším molárním mezenchymem. Obě tkáně byly pěstovány v předních komorách očí dospělých myší speciálního genetického kmene („nahé“ myši), které slouží jako neobvyklé, ale vhodné kultivační médium. Na rozdíl od očekávání reagoval kuřecí ústní epitel na myší ústní mezenchym vytvořením struktur připomínajících sklovinné orgány. V několika případech, z nichž jeden je znázorněn na obr. 5-8, se tyto kousky vyvinuly v pravé zuby. Genom alespoň jednoho druhu ptáka tak stále obsahuje genetickou informaci, která umožňuje kuřecímu ústnímu epitelu úspěšně se účastnit sekvenčních interakcí potřebných pro morfogenezi zubů a syntézu skloviny. Ztráta zubů u ptáků proto může být považována za důsledek změny ve vývojovém programu jejich mezenchymu, která vedla ke ztrátě počátečních fází tohoto procesu.
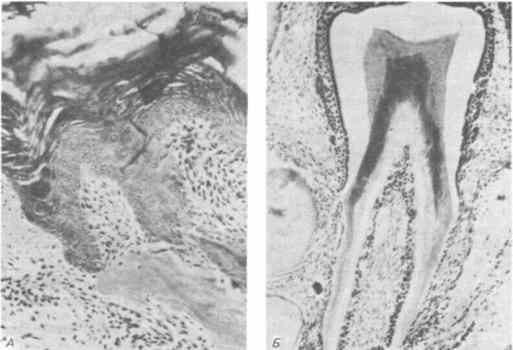
Rýže. 5-8. Hybridní zub vzniklý společným růstem ústního epitelu kuřecího embrya a zubního mezenchymu myši. A. Orální epitel kuřecího embrya, tvořící primitivní sklovinový orgán, který obsahuje mezenchym myšího embrya. B. Dobře vyvinutý hybridní zub (Kollar, Fisher, 1980).
Redukce fibuly u ptáků byla také důsledkem změny vývojového programu, nikoli ztráty genetické informace potřebné k vytvoření kosti. Archaeopteryx měl plně vyvinutou fibulu s kloubními plochami na obou koncích; naproti tomu u moderních ptáků (obr. 5-9) je fibula pouze kostěným výběžkem ležícím podél holenní kosti a srostlým s ní. Komplexní studii vztahů mezi stehenními kostmi u kuřecího embrya během vývoje provedl A. Hampe. Hampe provedl řadu experimentů, ve kterých byly různé oblasti vyvíjejícího se pupenu končetiny označeny částicemi dřevěného uhlí, které byly do nich vstříknuty. Umístění těchto značek ve vyvinuté končetině umožnilo Hampemu zmapovat potenciální oblasti pupenu končetiny (obr. 5-9). Hlavní tyto oblasti odpovídají akumulacím mezenchymálních buněk určených k vytvoření stehenní kosti, holenní kosti, fibuly, metatarzu a tarzu. Ampe provedl tři typy experimentů, které ho přesvědčily, že zmenšená velikost fibuly u moderních ptáků je výsledkem zachycení buněk patřících do prospektivní oblasti fibuly prospektivní oblastí holenní kosti.
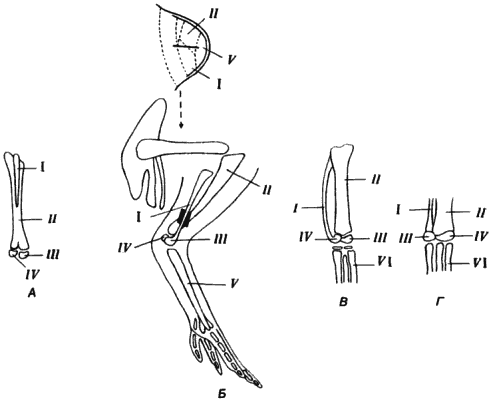
Rýže. 5-9. Atavismus projevující se ve vývoji nohy u kuřete s experimentální modifikací prospektivní oblasti kostní tkáně v pupenu končetiny. A. Normální kuřecí stehno. B. Zavedení slídové vločky do pupenu končetiny, mezi potenciální oblasti fibuly (I) a holenní kosti (II), a výsledná noha se zvětšenou fibulou nesoucí kloubní plošku na distálním konci. V. Projev atavismu ve struktuře kuřecí nohy. G. Dolní končetina Archaeopteryxe. III — fibulare; IV – holenní kost; V — tarsus; VI – metatarzu (Hampe, 1959, 1960).
První Ampéův experiment spočíval v odstranění nebo přidání mezenchymálních buněk do pupenu končetiny. Když byly buňky odstraněny z obou holenních kostí, konkurence mezi nimi se zvýšila a fibula se vůbec nevyvinula. Pokud byly mezenchymální buňky přidány do pupenu končetiny, tibie se nezměnila a fibula dosáhla stejné velikosti jako ona. Konkurenci bylo možné potlačit i jiným způsobem: otočení prospektivní oblasti tibie o 90° vedlo k růstu tibie a fibuly v různých směrech, takže konkurence mezi nimi se stala nemožnou. V tomto případě opět fibula dosáhla své plné délky. Tyto experimenty jasně odhalily konkurenci mezi prospektivními oblastmi obou holenních kostí; nejvýraznější a nejzajímavější informace o evolučních změnách, které se jich zúčastnily, však poskytl třetí experiment.
Ampé opatrně umístil malý kousek slídy mezi dvě perspektivní oblasti pupenu končetiny, což zabránilo pronikání předpokládaných buněk fibuly do oblasti holenní kosti. Získaný neočekávaný výsledek je znázorněn na obr. 5-9: ve vytvořené končetině fibula nejen dosáhla své „plné délky“, ale také nesla na svém distálním konci kloubní plošku. Tvar fibuly a její interakce se dvěma malými tarzálními kostmi, tibiale a fibulare, se velmi blížily homologním strukturám nohy Archaeopteryxe. U dospělého kuřete jsou tyto dvě kosti zcela srostlé s holenní kostí. Ampéovo oddělení perspektivních oblastí tibie a fibuly vedlo nejen ke vzniku delší fibuly, ale také k obnovení starodávného typu vývoje, který byl po dlouhou dobu potlačen. Geny, které určovaly tvar nohou u Archaeopteryxe, jsou u kuřete stále přítomny, ale v důsledku modifikací morfogenetických interakcí ztratily tyto geny schopnost být exprimovány, alespoň stejným způsobem jako dříve.
Příklady regulačních změn spojených se ztrátou zubů a redukcí lýtkové kosti u ptáků poskytují vhled do způsobů, jakými regulační mechanismy ovlivňující buněčné interakce mohou vést k evolučním změnám v morfologii. Oba příklady však trpí tím, že oba zahrnují redukci struktury. Pokud by evoluce usilovala o nějakou morfologickou nirvánu, bylo by to v pořádku, ale morfologická evoluce zahrnuje také modifikace struktur a někdy i vznik nových struktur. Bohužel případy, kdy evoluce vede ke ztrátě struktur (jako ve dvou již diskutovaných příkladech a v dalších, jako je ztráta očí u mexické jeskynní ryby studované Cahnem a Sadogluem, kteří prokázali geneticky podmíněné snížení schopnosti sítnice vyvolat vývoj čočky u těchto ryb), jsou přístupnější experimentální analýze než případy, kdy jsou struktury získány nebo modifikovány. Velkolepé transformace integumentů, ke kterým došlo u plazů, ptáků a savců, zahrnovaly komplexní změny v regulačních genových systémech a morfogenetické procesy, které jsou základem vývoje šupin, peří a srsti, vyžadují studium. Ačkoli je regulace morfogeneze v těchto systémech stále špatně pochopena, obecné obrysy evolučních změn v příslušných regulačních systémech se začínají objevovat z práce P. Sengela a jeho spolupracovníků, zejména Dhouaillyho.
Kůže se skládá ze dvou vrstev: horní vrstvy, epidermis, která pochází z ektodermu embrya, a podkladové vrstvy, dermis, která pochází z mezodermu. Vývoj šupin nebo jiných epidermálních struktur závisí na indukčních signálech z dermis.
Předpokládá se, že peří a srst se vyvinuly ze šupin plazů. Všechny tyto struktury se skládají z proteinů patřících do stejné rodiny keratinů. Úzká homologie mezi šupinami a peřím je naznačena někdy pozorovanou přeměnou špiček šupin pokrývajících nohy ptáků v peří.
Kachní peří se svou morfologií velmi liší od kuřecího peří. Sengel a jeho kolegové zkoumali zdroj morfogenetických informací nezbytných pro vývoj peří kombinací rudimentů dermis a epidermis odebraných z různých druhů. Ukázalo se, že morfogeneze peří je určena dermis. Obecná architektura peří, jejich velikost a počet ostnů odpovídaly druhu, ze kterého byla dermis odebrána. Epidermis určovala pouze tvar háčkových buněk. Dermis také určovala povahu rozložení peří. Jak ukázaly další experimenty s kuřecími embryi, ve kterých byla dermis kombinována s epidermis z dorzální oblasti, která dává vzniknout peří, a s epidermis z oblasti tarzu, která dává vzniknout charakteristickým velkým šupinám, je tento druh určující role dermis v morfogenezi rozšířený. Zda by epidermis byla určena k tvorbě rudimentů peří nebo velkých šupin, vždy záviselo na typu morfogeneze charakteristické pro část embrya, ze které byla dermis odebrána. Například dorzální epidermis, která obvykle tvoří peří, v kombinaci s dermis z oblasti tarsu vytvořila velké šupiny.
Šupiny i peří se skládají z keratinů, ale značně se liší v souboru keratinů, které obsahují. Při kombinaci dorzální a tarsometatarzální dermis a epidermis kuřecího embrya má epidermis, bez ohledu na to, odkud byla odebrána, možnost volby genu keratinu, který bude exprimován, a to v dermis. Uspořádání struktur tvořených epidermis, jejich morfologická specifičnost a exprese genů keratinu jsou tedy určeny indukčními signály přicházejícími z dermis. Zůstává však otevřená otázka, jak se tyto informační signály měnily během evoluce vyšších obratlovců.
Heterospecifické kombinace jsou možné nejen v rámci jedné třídy, ale i mezi třídami. Dhuayi provedl pozoruhodnou sérii experimentů kombinujících dermis a epidermis ještěrek, kuřete a myši v různých kombinacích. Jak tyto experimenty ukázaly, indukce se skládá ze dvou fází a tyto fáze se liší stupněm specificity. Některé z Dhuayiho výsledků jsou znázorněny na obr. 5-10. Reakce epidermis ještěrky na dermis kuřete nebo myši je obzvláště poučná. Epidermis ještěrky reaguje na tarsometatarzální dermis tvorbou velkých šupin, které jsou vzhledem a uspořádáním nerozeznatelné od šupin na tarzu ptáků. Epidermis ještěrky také reaguje na dorzální dermis kuřete, která u kuřat určuje rozložení a morfologii peří. Epidermis ještěrky však nedokáže interpretovat specifické signály, které způsobují, že kuřata produkují peří. Místo peří produkuje epidermis ještěrky nedostatečně vyvinuté šupiny uspořádané do šestiúhelníkového vzoru typického pro peří. Podobně epidermis ještěrky, když se spojí s dermis myši, vytváří nedostatečně vyvinuté šupiny uspořádané do typického vzoru srsti místo chlupů. Podobné vzorce jsou pozorovány i při kombinování tkání kuřat a myší. Umístění epidermálních struktur a raná stádia jejich diferenciace jsou řízeny regulačními látkami v dermis, které se v průběhu evoluce zachovaly takovým způsobem, že epidermální buňky patřící zástupcům jiných tříd jsou citlivé na jejich účinky. Pro úplnou diferenciaci základů šupin, peří nebo srsti jsou samozřejmě nezbytné signály specifické pro zvířata dané třídy.
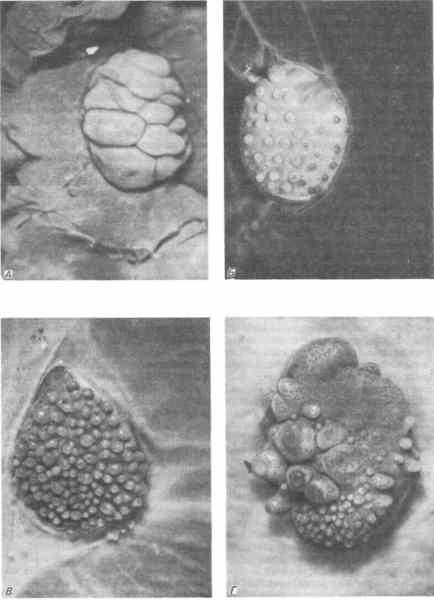
Rýže. 5-10. Morfologické struktury tvořené kombinacemi epidermis ještěrky, která normálně produkuje řady malých šupin, s dermis zástupců jiných tříd obratlovců v orgánových kulturách. A. Z kuřecí kůže v oblasti tarzu se tvoří šupiny, uspořádané jako na tarzu kuřete. B. S dorzální dermis kuřete se netvoří peří, ale nedostatečně vyvinuté šupiny, uspořádané do šestiúhelníkového vzoru typického pro peří. V. S dorzální dermis myši se tvoří šupiny, jejichž uspořádání odpovídá uspořádání primárních vlasových folikulů. G. V dermis myši z oblasti horního rtu se tvoří velké šupiny, uspořádané v souladu s typickým uspořádáním vibris a obklopené malými šupinami uspořádanými podobně jako vlasové folikuly srsti (Dhouailly, Sengel, 1973).
Více než 800 000 knih a audioknih!
Dostat 2 měsíce předplatného Litres jako dárek a užívejte si neomezené čtení